空间TCR文献分享--单细胞TCR映射揭示了头颈癌中空间协调的T细胞状态
原创
空间TCR文献分享--单细胞TCR映射揭示了头颈癌中空间协调的T细胞状态
原创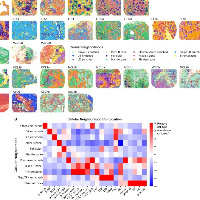
追风少年i
发布于 2026-05-13 10:08:32
发布于 2026-05-13 10:08:32
作者,Evil Genius
生活到处都是坑,越来越觉得,结婚真的是沉重的负担,如果所在城市没有房子,给孩子上的户口居然是集体户(我在太原),怪不得都不想结婚了,根本没办法结婚。
我现在都怀疑自己,跟人家姑娘结婚,是不是把媳妇和孩子都给坑了,先天就落后于人,瞬间觉得努力根本没啥意义。
看来结婚是有钱人的游戏。
今天我们分享文献,空间TCR,xenium的探针设计。

知识积累
肿瘤中存在肿瘤特异性T细胞(尤其在HPV阳性患者中),但其浸润与治疗成功之间的脱节表明,对有效抗肿瘤免疫的驱动因素仍缺乏理解。
现有矛盾与知识空白:
CD8+ T细胞虽是免疫检查点阻断(ICB)的靶点,但其存在与标准治疗后的良好预后相关,却与ICB应答无一致关联。
新证据表明,T细胞在肿瘤微环境(TME)中的空间组织是决定免疫治疗效果的关键。
但多数研究缺乏抗原特异性信息,因为人肿瘤中有大量“旁观者”(非癌症特异性)T细胞浸润。
核心内容
肿瘤浸润淋巴细胞(TILs) 的表型和TCR库与循环T细胞显著不同,仅靠血液分析无法反映瘤内免疫的复杂性。
空间异质性:即使临床和免疫学特征相似的肿瘤,其T细胞空间结构也存在显著差异,空间组织是一个独立且关键的免疫行为层面。
表型与位置的关联:
祖细胞样和记忆样T细胞定位于免疫富集区域。
耗竭T细胞则弥散分布于整个肿瘤中。
若没有克隆分辨率,这些空间模式可能被误读为特异性差异(如旁观者 vs 肿瘤特异性克隆)。
同一T细胞克隆的多样性:
携带相同TCR的T细胞表现出显著的转录组异质性,可同时包含干性样和耗竭样状态。
它们广泛分布于整个TME中。
抗原特异性的影响:
与旁观T细胞相比,肿瘤特异性克隆具有不同的邻近细胞类型和基因表达谱。
T细胞的分化不仅取决于克隆身份,更由抗原特异性与局部微环境信号的动态相互作用决定。
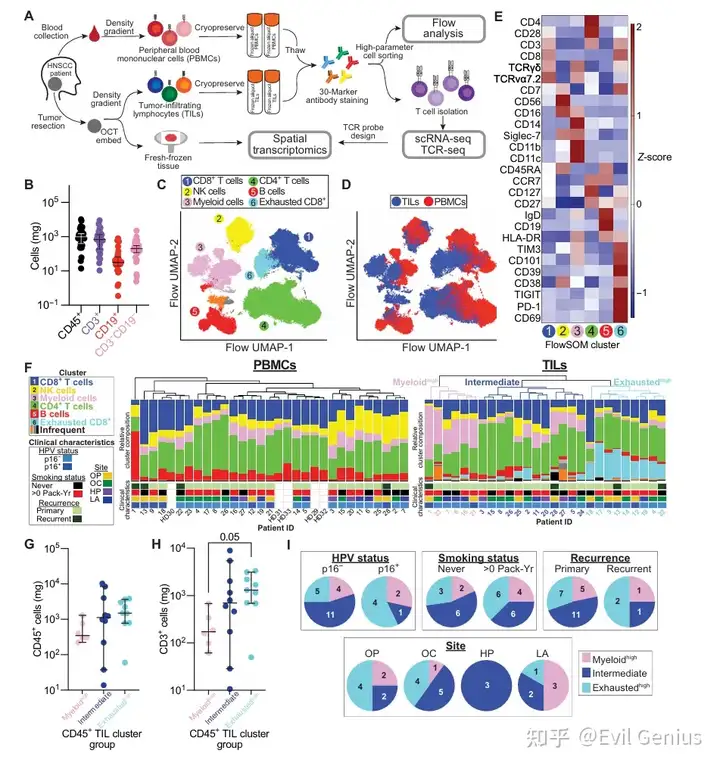
结果1、流式细胞术揭示了HNSCC中细胞组分特异性及患者间免疫异质性
在14个月内,前瞻性收集了27例HNSCC患者的配对肿瘤和血液样本。
部分肿瘤用于空间转录组学,分离外周血单个核细胞(PBMC)和肿瘤浸润淋巴细胞(TIL)并冻存。
使用31色荧光抗体对配对样本及5名健康供者的PBMC进行光谱流式细胞术分析,实现高维免疫 profiling 并分选活T细胞用于后续单细胞测序。
分析内容
免疫组成特征:
T细胞是多数肿瘤中的主要免疫细胞群,但患者间差异大,与p16状态、肿瘤部位、吸烟史、复发等临床特征无显著相关。
通过无监督聚类鉴定出9个免疫细胞群体,PBMC与TIL的免疫亚群丰度存在明显差异:
血液中富集NK细胞和B细胞。
TIL中富集髓系细胞和耗竭CD8+ T细胞,与TME支持功能失调/免疫抑制细胞的结论一致。
患者间免疫异质性(三个CD45+流式分组):
髓系高组(myeloidhigh):富集髓系细胞,CD45+和CD3+免疫细胞绝对数量较低。
耗竭高组(exhaustedhigh):富集耗竭CD8+ T细胞,常见于p16阳性、吸烟者、复发肿瘤、口咽或口腔肿瘤患者。
中间组:髓系和耗竭CD8+ T细胞频率较低,CD4+ T细胞和/或非耗竭CD8+ T细胞相对增多。
T细胞亚群分析:
PBMC中富集初始T细胞;TIL中富集调节性T细胞(Treg)、耗竭CD8+ T细胞和CD8+ 记忆样细胞。
耗竭高组中Treg和耗竭CD8+ T细胞最为丰富,进一步确认了各组独特的免疫格局。
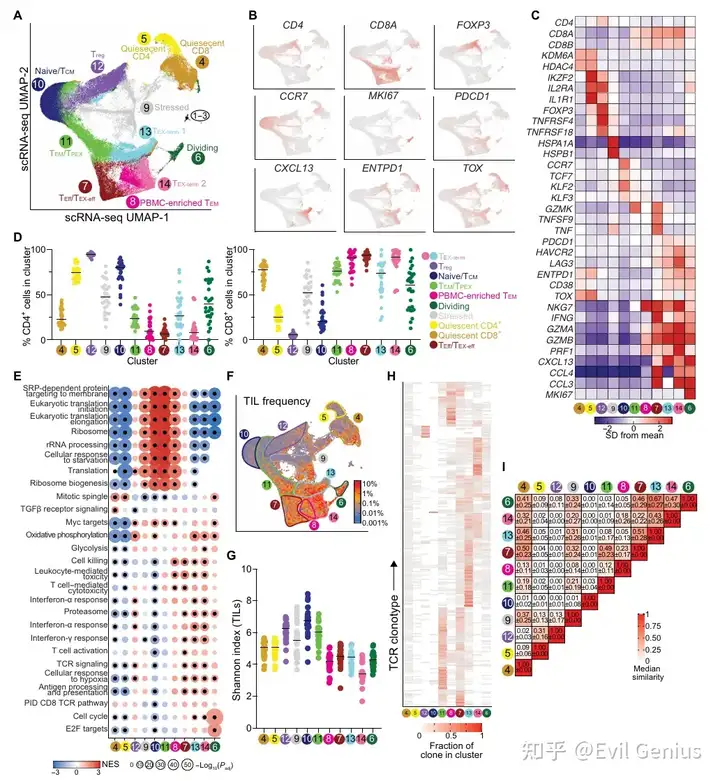
结果2、单细胞RNA测序定义了转录和克隆上不同的肿瘤浸润T细胞状态
克隆扩增与多样性:
耗竭克隆是扩增最明显的T细胞群体,克隆多样性最低。
利用“TEX相关”分析识别与耗竭T细胞共享TCR的非耗竭细胞,发现多数为分裂或效应细胞(中间耗竭状态),少数为表达TCF7的细胞(含效应记忆样和干细胞样耗竭前体TPEX)。TPEX细胞稀少且分散在记忆cluster中,未形成独立群体。
N/TCM细胞TCR多样性最高,Treg和效应记忆T细胞(TEM/TPEX)中等,分裂细胞多样性低(提示选择性进入细胞周期)。
TCR重叠分析:
最大的克隆共享出现在两个耗竭cluster之间,以及记忆样与效应cluster之间。
Treg细胞和PBMC富集的TEM细胞(非肿瘤特异性CD8 T细胞)与其他cluster的TCR重叠极少,提示它们与瘤内T细胞主要分化轨迹相对独立。
Treg细胞异质性:
Treg可细分为三个亚群(Treg1→Treg3),形成从CCR7/TCF7富集表型向高表达免疫抑制基因(CCR8, FOXP3, IL10)的连续分化轴。
伴随转录演进,克隆多样性下降,Treg2与Treg3之间高克隆重叠支持共享分化路径,而Treg1与其他亚群相似性低,提示其更具可塑性或尚未完全定型。
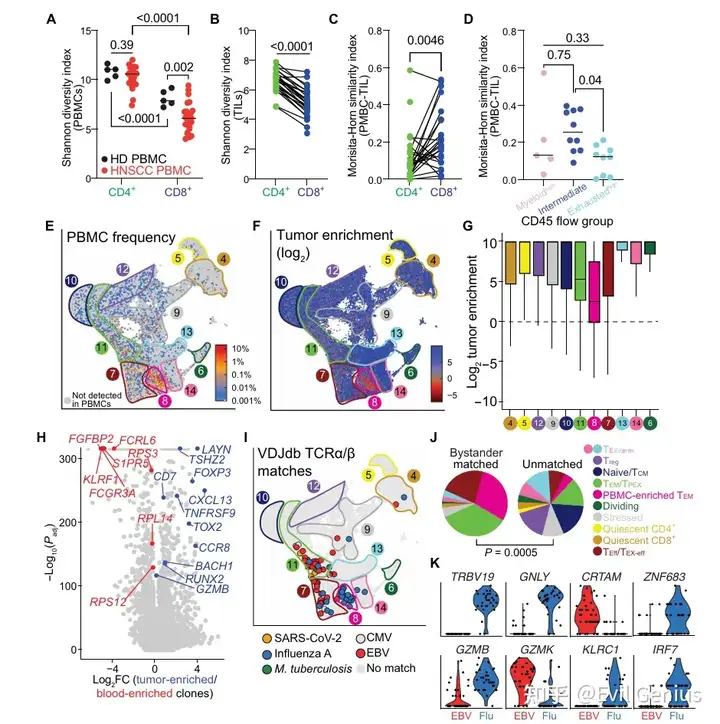
结果3、循环和肿瘤浸润的T细胞表现出不同的克隆性和表型景观
比较头颈鳞癌患者血液循环(PBMC)与肿瘤浸润(TIL)中T细胞的克隆结构和表型差异,并分析非常规T细胞(γδ T、MAIT细胞)的特征。
区室特异性克隆格局:
无论是患者还是健康供者,PBMC中CD8+ T细胞的克隆扩增程度均高于CD4+ T细胞,其中HNSCC患者的CD8+ T细胞多样性最低。
TIL中CD8+ T细胞的多样性同样低于CD4+ T细胞。
PBMC与TIL之间的克隆重叠有限,CD8+ T细胞重叠略高于CD4+ T细胞。耗竭高组的克隆重叠更低,提示瘤内耗竭增加与血液循环TCR库的趋异程度加大相关。
循环来源与肿瘤来源T细胞的转录差异:
血液循环中丰度高的TCR主要定位于PBMC富集cluster(Pb-enr cluster,即cluster 8),该cluster的肿瘤富集度最低。
肿瘤富集克隆上调调节、效应及耗竭相关基因(如FOXP3、GZMB、CXCL13)。
血液富集克隆高表达核糖体蛋白(RPS3、RPL14)、迁移相关基因(S1PR5)及NK样基因(FCGR3A)。
FGFBP2是血液富集克隆的强标志物,可能反映T细胞从循环向TME的浸润能力(独立于抗原特异性)。
抗原特异性分析(基于VDJdb数据库):
识别已知微生物抗原(如EBV、甲流病毒)的TCR主要存在于PBMC富集TEM、Teff/Tex-eff、TEM/TPEX等cluster中。
EBV和甲流特异性T细胞分别定位到不同cluster并呈现不同基因表达程序,提示病原体特异性免疫印记可能导致旁观者T细胞的异质性,进而影响TME整体免疫格局。
非常规T细胞(γδ T与MAIT细胞):
γδ T细胞:主要分布于终末耗竭、TEM/TPEX、PBMC富集TEM等cluster;表达效应/NK样转录特征(NCAM1、GZMB、KLR家族);在血液循环中比肿瘤中更丰富;多数瘤内γδ T细胞为CD4−CD8−,但仍主要定位于CD8富集cluster。
MAIT细胞:广泛分布于所有T细胞cluster;表达典型标志物(如KLRB1/CD161),下调干性相关基因(如SELL/CD62L);肿瘤富集度较低(可能特异性针对微生物而非肿瘤抗原);HNSCC患者循环MAIT细胞较健康供者减少,与其他癌症研究一致。
血液循环与肿瘤浸润T细胞在克隆组成、转录表型和抗原特异性上存在显著差异;非常规T细胞亚群(γδ T、MAIT)在TME中具有独特分布和表型特征;外周血TCR谱不能完全反映瘤内T细胞状态,且病原体免疫记忆可能塑造肿瘤免疫微环境。
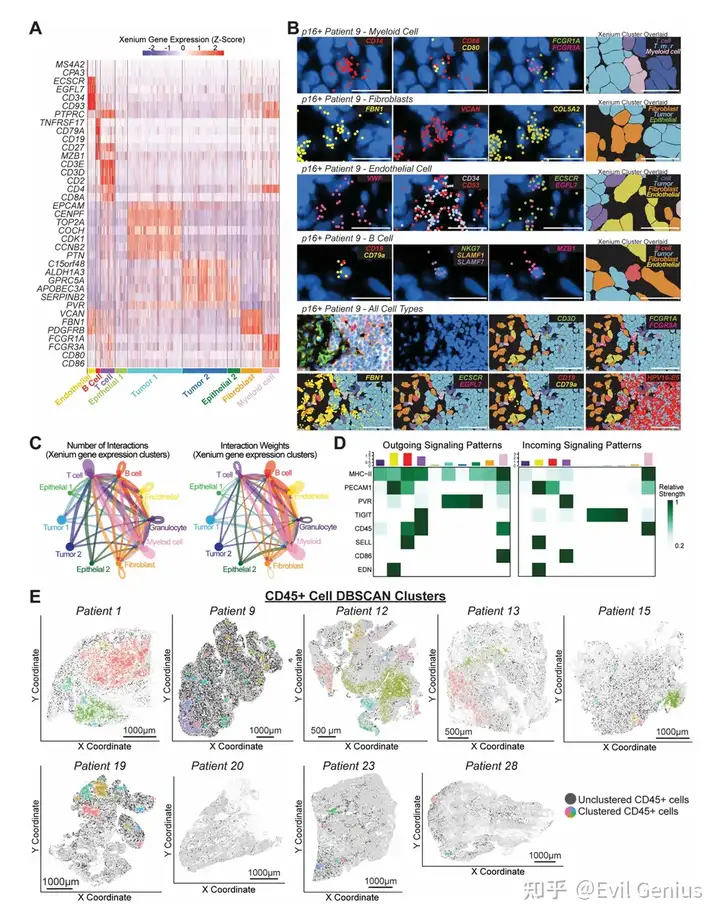
结果4、空间转录组学在单细胞分辨率下定义了HNSCC的细胞景观
肿瘤切片进行了Xenium空间转录组学分析。
单细胞精度识别:
能够精确识别细胞类型,如共表达CD3D/CD8A和TRAC/TRBC2的CD8+ αβ T细胞。
效应潜能标志:GZMA/GZMB;耗竭表型标志:ENTPD1、HAVCR2、TOX。
可观察到抑制性受体-配体对(如TIGIT与PVR)的潜在免疫调控互作。
可区分Treg细胞(FOXP3+IL2RA+)和记忆/干细胞样T细胞(TCF7、CCR7、IL7R)。
细胞聚类与组成:
通过UMAP和聚类分析鉴定出10个不同的基因表达cluster,包括:T细胞、B细胞、髓系细胞、内皮细胞、成纤维细胞、粒细胞以及四个上皮亚群(两种恶性/肿瘤上皮,两种正常上皮)。
所有cluster在每位患者中均存在,但相对比例各异,反映了患者间TME组成的异质性。
9/10的患者以肿瘤cluster细胞为主;患者1(合并慢性淋巴细胞白血病)表现为高比例B细胞、极低上皮细胞的不典型特征。
cluster组成与其他临床因素无明确相关性。
HPV+肿瘤验证:
使用定制Xenium探针靶向HPV16基因产物,在HPV+肿瘤中可选择性表达,有效鉴定肿瘤细胞。
验证与准确性:
将基因表达cluster叠加到组织图像上,确认谱系定义转录本定位于相应细胞。
谱系特异性标志物相对局限于对应细胞类型,支持在复杂HNSCC TME中细胞类型分类和细胞分割的准确性。
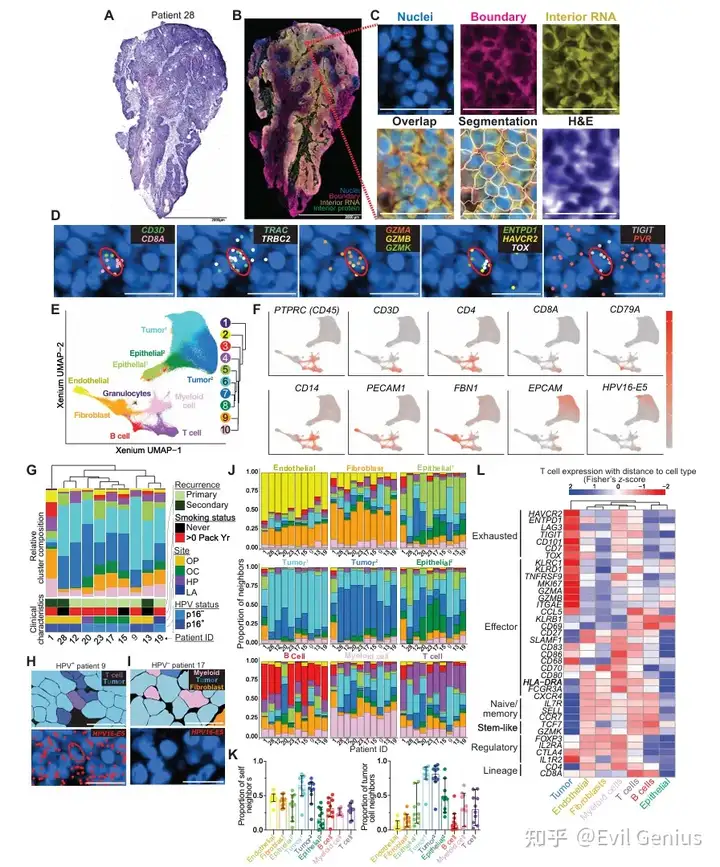
结果5、细胞类型在HNSCC肿瘤微环境(TME)中表现出不同的空间相互作用。
不同细胞类型的空间邻域特征:
内皮细胞和肿瘤细胞:最常与自身同类细胞相邻,分别形成血管区和上皮区。
成纤维细胞:几乎与所有细胞类型相邻,也彼此相邻,形成广泛分布于整个肿瘤的成纤维细胞富集区。
T细胞和髓系细胞:相较于其他免疫或基质细胞,更接近肿瘤细胞,提示它们优先定位于恶性区域,而非外周或血管区域。
B细胞:主要彼此相邻,与肿瘤细胞接触有限。
细胞间通讯预测(CellChat分析):
T细胞与髓系细胞之间预测的相互作用最强,凸显了它们在TME通讯中的核心地位。
关键通路包括:肿瘤与T细胞之间的PVR-TIGIT(免疫抑制信号)、髓系向T细胞的CD86介导信号(共刺激轴)。
T细胞转录状态与空间位置的关联:
靠近肿瘤细胞和髓系细胞的T细胞:高表达耗竭相关基因(ENTPD1、HAVCR2、LAG3、PDCD1)、效应分子(GZMA、GZMB)和活化标志物(HLA-DRA、SLAMF1),提示在肿瘤抗原刺激下被激活。
靠近正常上皮细胞的T细胞:相比邻近恶性上皮细胞的T细胞,耗竭标志物表达降低,但仍表达组织驻留记忆标志物(CD69、ITGAE/CD103)。
远离肿瘤/髓系细胞、靠近B细胞/内皮细胞/其他T细胞的T细胞:富集初始和记忆相关转录本(IL7R、SELL、CCR7、GZMK)。
靠近成纤维细胞、内皮细胞和髓系细胞的T细胞:调节性标志物(FOXP3、IL2RA)表达最高。
结论:
HNSCC肿瘤微环境中,T细胞占据多样化的空间背景:
肿瘤/髓系细胞附近:呈现激活和耗竭特征;
成纤维/内皮/髓系细胞附近:呈现调节性特征;
B细胞/内皮/成纤维/其他T细胞富集区域:呈现更初始或记忆样特征。
这表明T细胞的功能状态与其微环境空间位置密切相关。
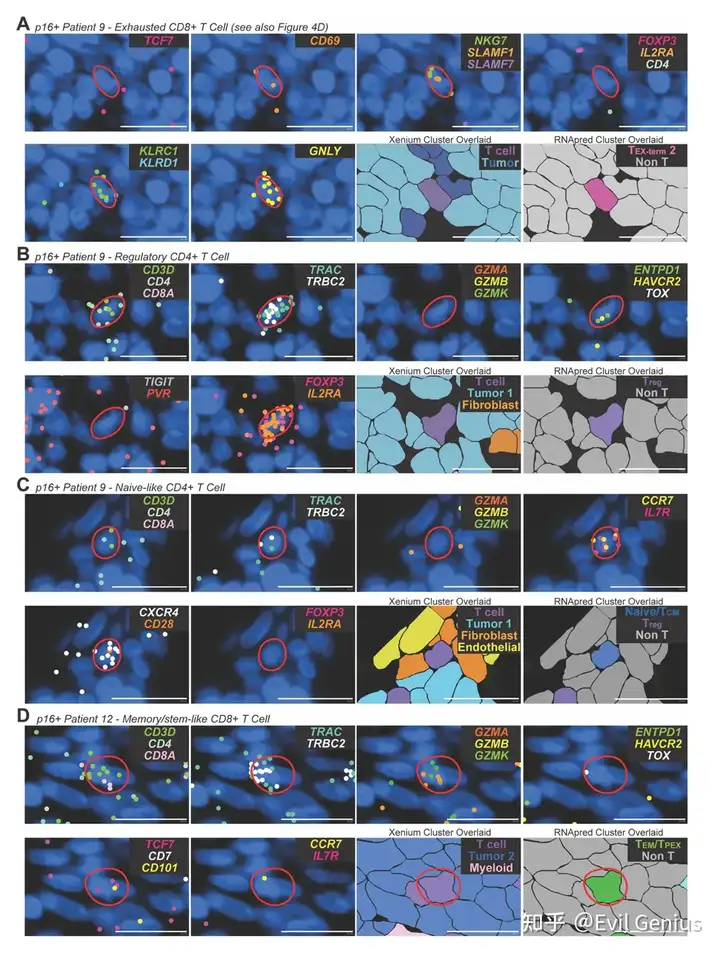
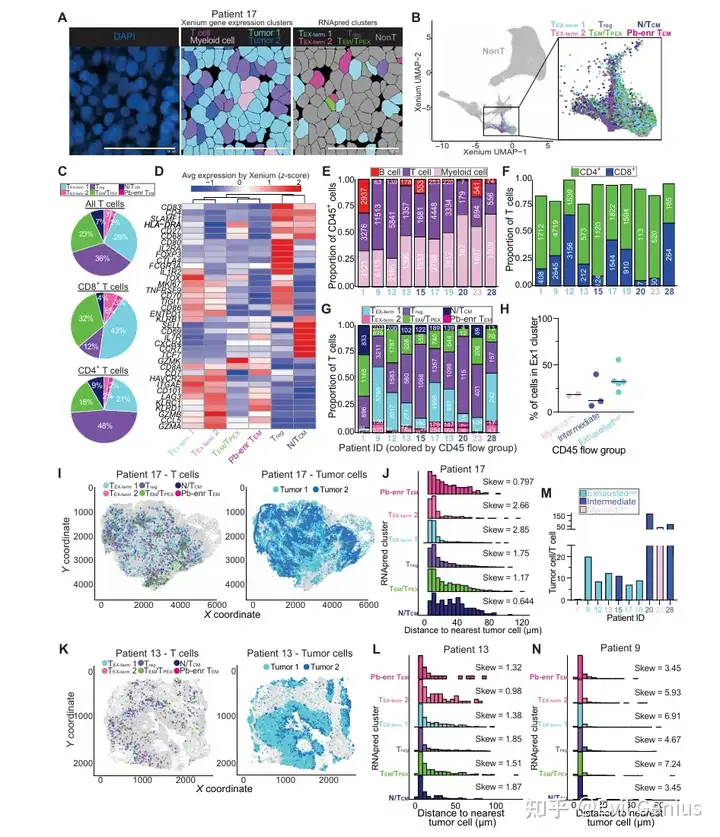
结果6、scRNA-seq与Xenium的整合优化了空间分辨T细胞的表型分类
Xenium空间转录组学的基因覆盖范围有限,难以精确分类T细胞亚群。为此,利用患者匹配的scRNA-seq T细胞图谱,通过随机森林分类器将scRNA-seq中的T细胞亚群标签迁移到Xenium数据中,从而实现空间T细胞的表型精细分类。
主要方法与验证:
分类器基于两个数据集共有的基因进行训练,排除了分裂细胞和低转录细胞(这些状态可能是暂时的,且难以与技术变异区分)。
预测得到的RNA-seq cluster标签(RNApred)被赋予每个Xenium T细胞。
模型在保留的scRNA-seq数据上评估,总体准确率为75.6%。
主要发现:
空间T细胞组成:
各样本中,Xenium检测到的T细胞主要为终末耗竭T细胞(TEX-term)、效应记忆/干性样耗竭前体T细胞(TEM/TPEX)和调节性T细胞(Treg)。
CD8+ T细胞以TEX-term和TEM/TPEX为主;CD4+ T细胞中近半数为Treg细胞。
RNA pred cluster的Xenium基因表达谱重现了scRNA-seq中的关键转录特征(如TEX-term高表达HAVCR2、TOX;TEM/TPEX高表达GZMK;Treg表达FOXP3、IL1R2;N/TCM表达CCR7、IL7R、SELL)。
跨平台一致性:
流式细胞术定义的髓系高组中,Xenium也显示CD8+ T细胞含量及耗竭比例降低,验证了跨平台结果一致。
T细胞表型的空间分布:
典型患者(患者17)中,TEX-term细胞嵌入肿瘤密集区,而N/TCM、TEM/TPEX和Treg细胞富集于T细胞丰富区域。
距离计算证实:TEX-term细胞更倾向于靠近肿瘤细胞,而N/TCM、Treg和TEM/TPEX细胞则相对远离。
肿瘤微环境空间异质性的影响:
上述空间模式并非在所有患者中均成立:
免疫排斥型肿瘤(患者13)未见此相关性。
在肿瘤与T细胞比例高的患者中,几乎所有T细胞天然接近肿瘤细胞,空间关系分析缺乏动态范围。
这些发现强调了:看似在流式或scRNA-seq中相似的患者样本,其空间组织可能高度不同,空间背景能够揭示传统单细胞分析无法捕捉的生物学变异。
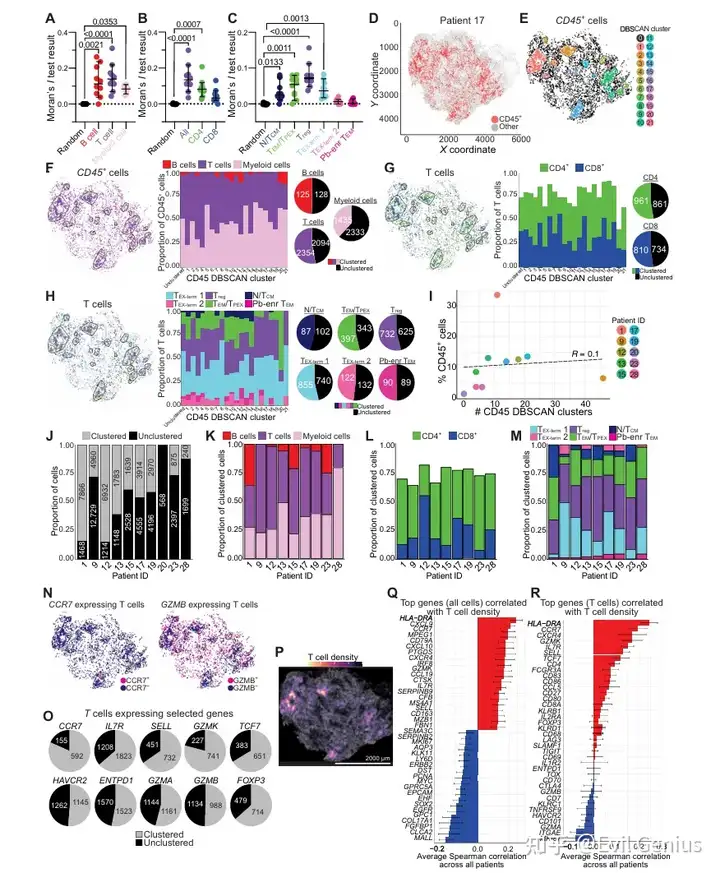
结果7、空间聚类识别出具有不同T细胞表型的有序免疫微环境
免疫细胞的空间自相关性(Moran's I分析):
免疫细胞整体呈现显著的空间聚集,与随机分布相比有显著差异。
在T细胞亚群中,CD4+ T细胞(而非CD8+ T细胞)表现出比随机分布更强的自相关性。
在RNApred定义的T细胞亚群中,Treg细胞和TEM/TPEX细胞的自我聚集性最强。这些趋势在不同患者中相对保守。
无监督密度聚类(DBSCAN)识别免疫聚集区:
DBSCAN成功识别出高免疫细胞密度区域。典型患者(患者17)中,大多数DBSCAN聚集区主要由T细胞和髓系细胞组成,T细胞优先位于聚集区内,而髓系细胞更多分散在聚集区外。
在CD45+聚集区中,CD4+ T细胞数量超过CD8+ T细胞;TEM/TPEX、Treg和TEX-term细胞在聚集区中富集。
不同患者的CD45+ DBSCAN聚集区数量各异,与总免疫浸润量和临床特征无关,反映了免疫组织的空间异质性。
跨患者保守的聚集模式:
尽管患者间存在差异,但整体队列中出现了保守的聚集模式:CD45+生态位持续富集T细胞和髓系细胞,包括CD4+ T细胞丰度高于CD8+ T细胞,以及TEX-term、Treg和TEM/TPEX亚群的优先定位。
不同T细胞状态标志物的空间分布差异:
初始/干性样标志物(CCR7、IL7R、SELL、TCF7)、记忆相关标志物(GZMK)和调节性标志物(FOXP3)主要位于CD45+ DBSCAN聚集区内。
效应和耗竭标志物(HAVCR2、ENTPD1、GZMA、GZMB)则更常见于聚集区外。
T细胞密度区域的特征基因富集:
高T细胞密度区域:富集初始/淋巴谱系相关基因(CCR7、CD79A)以及T细胞中的初始/记忆样基因(CCR7、SELL),反映淋巴样生态位。
低T细胞密度区域:富集肿瘤相关基因(MALL、SOX2)以及T细胞中的耗竭/驻留相关基因(HAVCR2、CD101、ITGAE),以肿瘤细胞为主。
总结一下
耗竭T细胞(TEX) 主要分散分布于肿瘤密集区。
非耗竭T细胞亚群(初始/干性样、记忆样、调节性)则集中分布于免疫密集区(免疫生态位)。
这些空间组织模式在不同患者中相对保守,揭示了TME中T细胞功能状态与空间位置之间的系统性能关联。
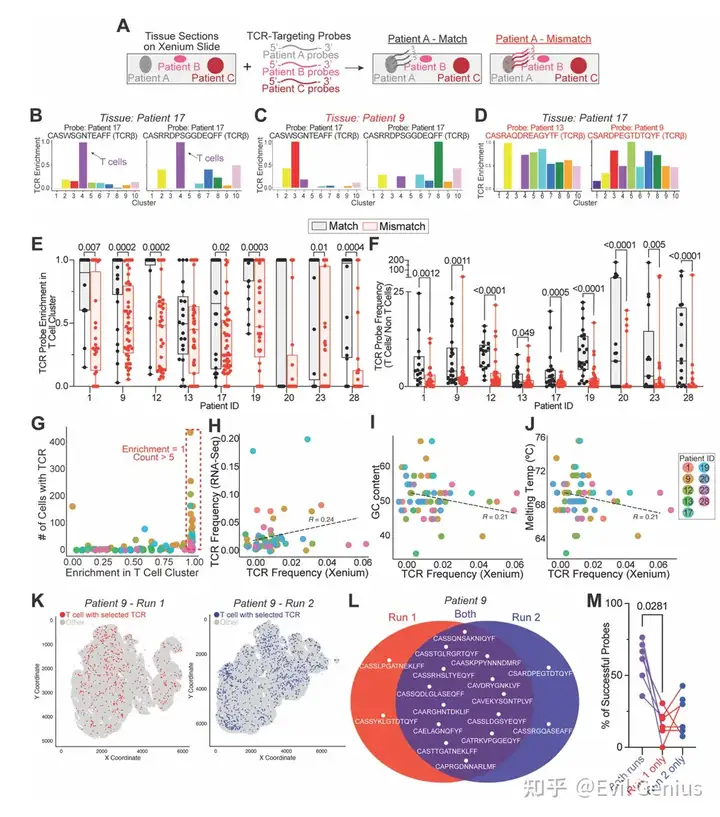
结果8、空间映射在单细胞分辨率下解析肿瘤浸润T细胞克隆个体
为了直接在原位评估克隆身份,研究者开发了一种策略:针对每位患者,设计定制的、患者特异性的TCR靶向探针(针对肿瘤浸润丰度较高的TCR可变区),在Xenium空间转录组学平台上实现单细胞分辨率下对单个T细胞克隆的空间定位。
技术验证:
每次Xenium实验包含至少三位不同患者的TCR探针,使得不匹配的样本-探针组合可作为内部阴性对照。
大多数探针仅在对应患者的组织T细胞中被特异性检测到,而在错配样本中无信号,证实了患者特异性和细胞类型特异性。
成功验证了61个探针,在8位患者的2000多个T细胞中检测到目标克隆。
scRNA-seq与Xenium中检测到的TCR频率呈中等程度相关。
检测效率与探针的GC含量和熔解温度无关。
在7例重复测定的样本中,6例在两个独立Xenium运行中均成功检测到TCR探针,且探针重叠显著,证明方法具有良好的可重复性。变异主要来源于不同切片区域的组织含量和克隆代表性差异。
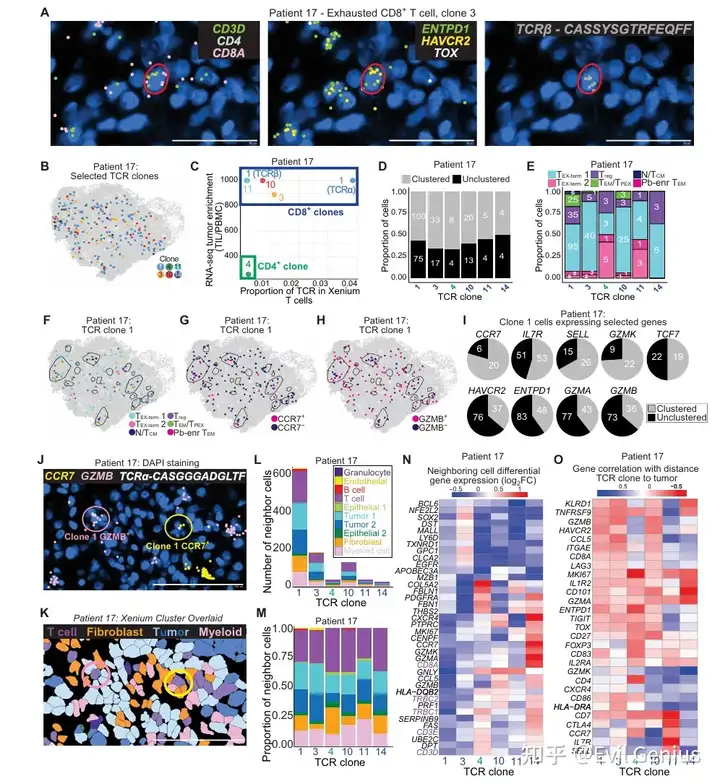
结果9、T细胞克隆的映射揭示了空间协调的功能状态
将单个T细胞克隆的空间分布与功能状态直接关联,探究克隆身份如何影响T细胞在肿瘤微环境中的空间组织和功能状态。
克隆的总体分布特征:
检测到6个肿瘤富集的TCR克隆,每个克隆均广泛分布于整个组织。
所有克隆均倾向于定位于CD45+ DBSCAN免疫聚集区内,且主要由终末耗竭T细胞(TEX-term)表型组成。
约10%的CD8+ T细胞被预测为Treg表型,这可能反映了分类器75.6%的准确率以及耗竭T细胞与Treg细胞在有限基因面板下的转录重叠(如均表达PDCD1、CTLA4、TIGIT等)。
同一克隆内的表型与空间异质性(以最丰富的CD8克隆1为例):
同一个克隆中,RNApred定义的不同亚群以及表达干性样/耗竭基因的细胞在空间上明显分离。
更具可塑性的状态(如表达CCR7的细胞)富集于CD45+免疫聚集区内;耗竭状态(如表达GZMB的细胞)则更分散。
关键实例:克隆1的两个邻近细胞——一个表达GZMB(位于免疫聚集区外,被肿瘤细胞包围),另一个表达CCR7(位于免疫聚集区内,邻近髓系细胞和成纤维细胞)。这直接证明了:共享同一TCR的细胞可以采取不同的表型并占据不同的组织微环境。
克隆间的空间差异:
不同克隆在邻近细胞类型上存在差异:
CD4克隆(克隆4):更常邻近成纤维细胞。
CD8克隆(克隆3):更常邻近肿瘤细胞(邻居富集NFE2L2、MALL)。
CD8克隆(克隆14):更常邻近其他T细胞(邻居富集CD3E、CD8A)。
基因表达与肿瘤距离的相关性也因克隆而异:
克隆1、3、10:靠近肿瘤细胞时,耗竭标志物表达增加。
CD4克隆:靠近肿瘤细胞时,FOXP3表达增加。
单个T细胞克隆内部存在显著的功能和空间多样性。
同一克隆的T细胞可以在不同微环境背景下分化为不同状态(可塑性/干性样 vs 耗竭),而非克隆身份单一决定命运。
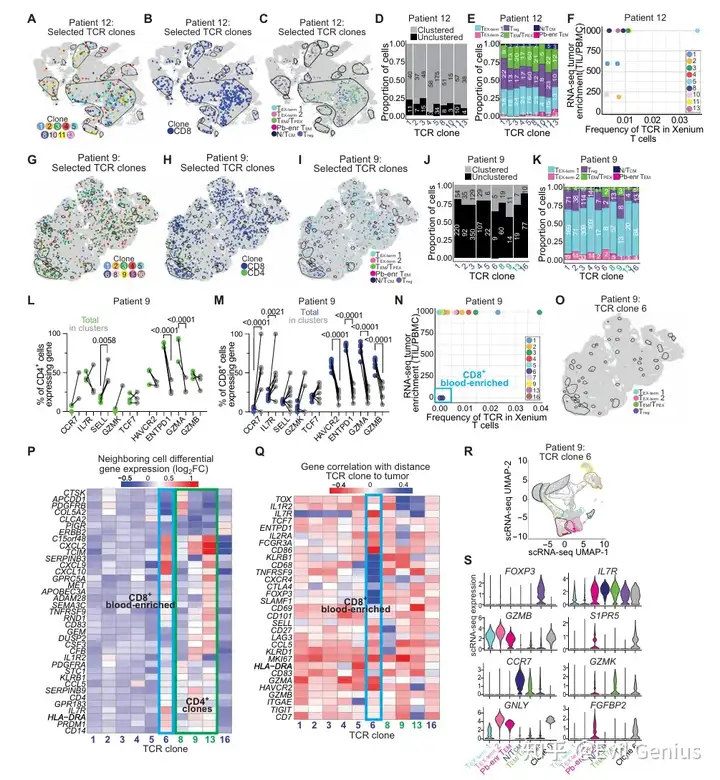
结果10、T细胞克隆在患者间显著不同,包括聚类、表型和肿瘤富集
比较两位临床特征相似(均为p16阳性、吸烟者、原发肿瘤、流式定义的耗竭高组)但空间免疫组织截然不同的患者(患者12:存在大的免疫生态位;患者9:免疫细胞弥散分布),探究T细胞克隆的空间分布、表型和肿瘤富集程度的差异。
T细胞克隆的空间分布、转录状态和肿瘤富集程度在不同患者间存在显著差异,定义了异质性的瘤内T细胞克隆结构。
这些差异无法仅通过流式细胞术、单细胞RNA测序或临床变量完全捕捉,凸显了空间分析在解析肿瘤免疫复杂性中的独特价值。
大的免疫生态位可能为干细胞样/可塑性T细胞(TEM/TPEX)提供支持性微环境,而弥散分布的患者则以终末耗竭T细胞为主。
生活很好,有你更好
原创声明:本文系作者授权腾讯云开发者社区发表,未经许可,不得转载。
如有侵权,请联系 cloudcommunity@tencent.com 删除。
原创声明:本文系作者授权腾讯云开发者社区发表,未经许可,不得转载。
如有侵权,请联系 cloudcommunity@tencent.com 删除。
评论
登录后参与评论
推荐阅读
目录
腾讯云开发者

Copyright © 2013 - 2026 Tencent Cloud. All Rights Reserved. 腾讯云 版权所有
深圳市腾讯计算机系统有限公司 ICP备案/许可证号:粤B2-20090059 ![]() 粤公网安备44030502008569号
粤公网安备44030502008569号
腾讯云计算(北京)有限责任公司 京ICP证150476号 | 京ICP备11018762号