全球生态研究如何确定植被生长季:126篇文献方法总结

全球生态研究如何确定植被生长季:126篇文献方法总结

疯狂学习GIS
发布于 2026-06-02 13:55:18
发布于 2026-06-02 13:55:18
本文介绍全球高水平生态环境遥感研究中,植被生长季的定义与筛选方法。

在植被生态学研究中,一个看似简单却极容易被忽视的前处理步骤是:如何只保留生长季数据?
如果我们要做(或者曾经做过)全球尺度的植被-气候关系分析,大概率会遇到(或者遇到过)这样的困扰:把全年NDVI和全年降水做相关性,发现干旱区的相关系数显著且漂亮——但细看才发现,冬季的裸土低值与冬季的微量降水恰好形成一组"伪相关"。非生长季的植被信号,比如积雪覆盖的反射率跳变、裸土背景噪声、休眠期近乎恒定的低值等,如果不加剔除,会像地毯下的灰尘一样被悄悄卷进拟合模型,污染你对生态系统真实响应的判断。
但如何定义"生长季",不同研究之间差异巨大,且往往缺乏充分讨论。很多论文在Methodology部分只用一两句话带过,我们很难从字里行间还原操作全貌。
本文基于我之前陆陆续续整合的Zotero + ima知识库,从方法论层面归纳了现有研究生筛选生长季数据的五类主要方法,并结合具体案例讨论各类方法的逻辑基础、适用范围与潜在盲区。
1 问题从何而来
要理解为什么生长季筛选如此重要,不妨先退一步看:植被遥感信号到底在"说"什么?
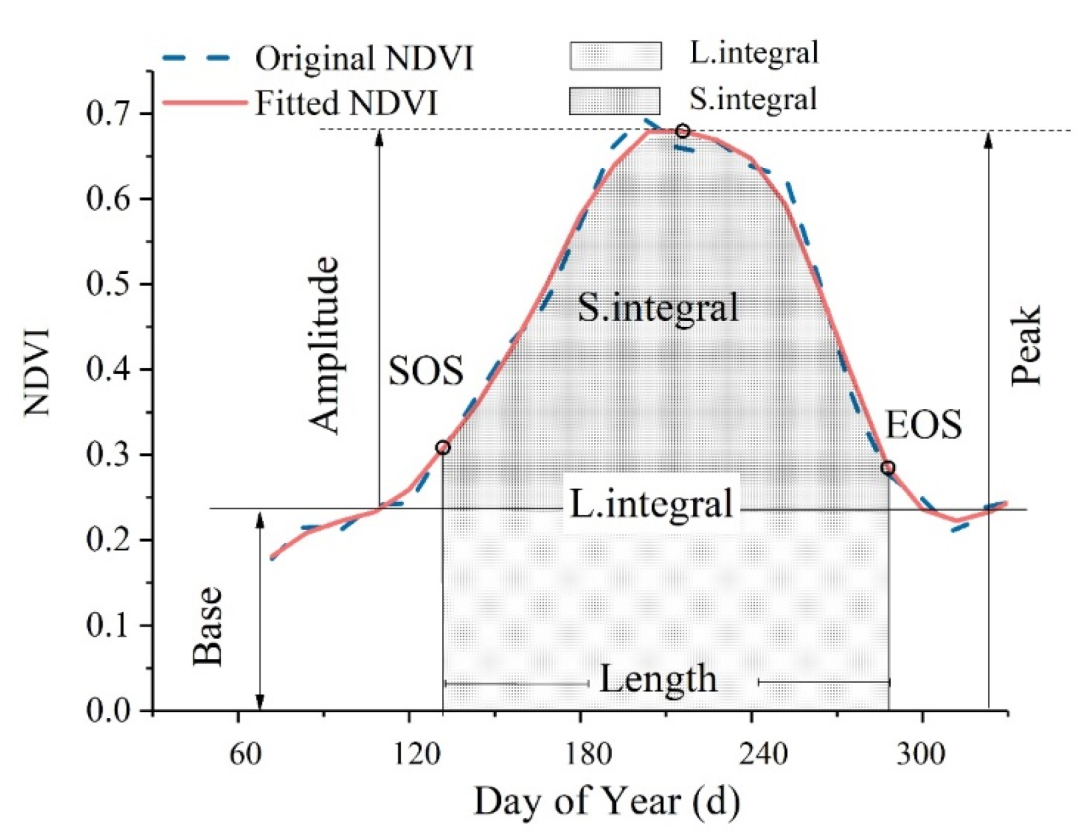
以最常用的NDVI为例,其月值序列中至少混合了三类信号:一是物候驱动的季节循环(春天萌发、夏季峰值、秋冬休眠),这是最大振幅的信号分量;二是干旱等极端事件造成的短期异常(通常比季节振幅小一个量级);三是长期趋势(如CO₂施肥效应导致的渐进绿化)。随便举个例子:我们的研究目标是"干旱对植被的影响",那真正想捕捉的恰恰是第二类,也就是那个被季节循环淹没的小信号。
这就构成了一个方法论上的两难:不剔除非生长季,噪声太大;剔除方法太粗糙,信号跟着一起丢。
打个不严谨但直观的比方:你在一段嘈杂的录音里寻找一个微弱的敲击声。如果你知道敲击声只出现在某个特定频段,把其他频段滤掉是最直接的做法。但如果你不知道这个频段的确切范围,或者这个范围本身每年都在漂移,你就需要在"滤太窄漏信号"和"滤太宽进噪声"之间权衡。
更麻烦的是,气候变化本身就在改变物候。过去三十年,北半球中高纬度的春季开始日期持续提前、秋季结束日期持续推迟。这意味着如果你用2000年的气候态来划定2020年的生长季,这批"固定"的月份区间可能已经系统性偏窄了——那些提前萌发的春季信号和延后衰老的秋季信号,恰恰是对气候变暖最敏感的时段,却没被纳入分析。
2 方法谱系
基于现有文献,生长季数据的筛选方法大致可归入五个基本类型。这当然不是严格的分类学,很多研究综合采用了多种手段;但从方法逻辑上看,每条路径代表了对"什么是生长季"这一根本问题的不同理解。

2.1 固定月份法:一刀切的理性与代价
这是最常见的方法族:直接指定几个月份作为生长季,其余月份的数据一概剔除。操作简单、逻辑透明,是所有方法中可复现性最高的。
但"固定"并不意味着"统一"。文献中至少存在三种变体:
全域统一月份是最原始的形式。例如Peng et al. (2013)在Nature上发表的昼夜不对称增温研究中,对北半球取4–10月;Ji and Peters (2003)针对美国北部大平原取5–10月;Deng et al. (2021)和Liang et al. (2026)对中东亚取4–10月。这种做法适用于研究区域集中在单一半球、纬度跨度不大的情况,但显然不适用于跨南北半球的全球分析。
一个值得单独提及的特例是Konings et al. (2017)的做法:仅取7–8月峰值NDVI来代表年际变化。理由是生长季峰值期间植被变异性最大,对气候信号的响应最敏感。这在逻辑上说得通,但代价是丢失了整个春季萌发和秋季衰老期间的信息,而这两个阶段恰恰可能是干旱影响植被的关键窗口;不过也不耽误,因为你只要在题目、开头说清楚,你分析的只是生长季峰值期间的规律即可。
按半球/纬度分区是在全球尺度不得不做的妥协。典型做法是按南北30°或赤道为界,南北半球分别取互为对跖的月份区间,例如最简单的取北半球4–9月、南半球10–3月;Xi and Yuan (2022)在上述分区基础上对热带保留全年数据;O and Park (2024)则以30°为界,30°S–30°N全年保留,高纬度分别取3–10月和9–4月。
这种分区策略在操作上合理,但30°这个阈值本身是有问题的——它大致对应副热带高压带的位置,与生态系统的物候边界并无直接关系。热带稀树草原有明显的干湿季交替,并不会因为地处赤道就全年均质生长。而且有的研究取30°,有的则是23.5°,到底哪一个更好?说不清的好吧。
固定月份+植被指数掩膜则是在月份基础上叠加第二道筛子。Wang et al. (2026)取5–10月后排除NDVI < 0.075的像素,Lu et al. (2026)取3–11月后排除生长季平均EVI < 0.1的像素。这种组合策略能有效剔除月份区间内仍存在裸土或极稀疏植被的像元,成本几乎为零,收益明确。
固定月份法的根本矛盾在于:它是一种空间上"硬截断"——既不能反映不同植被类型之间物候节律的巨大差异(温带落叶林和地中海灌丛的生长季月份几乎不重叠),也不能捕捉年际间生长季长度的变化。但在计算效率和可复现性方面,它仍然是最稳健的选择,尤其在大数据时代需要批量处理成千上万像元时。
2.2 植被指数阈值+温度双条件法:简单背后的校准难题
如果你的研究恰好避开了固定月份法的主要缺陷(需要跨半球、跨生物群系的分析),但又不想引入复杂的物候模型,那么植被指数阈值加温度条件的双门槛方案是一种自然的中间选择。
核心逻辑一目了然:温度达到生理下限+植被指数达到活性下限,双重条件同时满足才算生长季。这种做法最早由Piao et al. (2006)提出(NDVI > 0.2且月均温 > 0°C),此后被Wu et al. (2015)、Ding et al. (2020)、Wen et al. (2019)等多篇论文直接引用沿用。
Ding et al. (2020)还进一步将生长季细分为NDVI上升期(生长阶段)和下降期(衰老阶段),以区分不同物候阶段对气象条件的响应差异。这意味着作者关心的并非"是否为生长季"这个二元问题,而是生长季内部不同物候阶段的差异化响应。
温度阈值的选择也有变化。Li et al. (2022, 2023)将温度门槛从0°C提高至5°C,分别使用负值植被指数过滤和SIF多年平均季节循环值 > 0.2 mW m⁻² sr⁻¹ nm⁻¹作为植被活性判据。Li et al. (2023)特别提到,使用SIF的多年平均季节循环(而非当年动态值)是为了保留旱后植被异常信号——如果在旱年用当年SIF做阈值,异常低值本身会被错误剔除。这是一个极其关键但容易被忽视的细节:你的筛选条件本身可能和被筛选对象是信息共变的。
双条件法的阿喀琉斯之踵是阈值选择缺乏客观标准。NDVI > 0.1和NDVI > 0.2的差别,在稀树草原和灌木丛这类低覆盖度生态系统中,可能意味着保留或丢弃大量有效数据。温度阈值也同样:0°C大致对应生理零度,排除冻土;5°C更保守,对应多数C3植物的活跃光合起点。但0°C在温带可能包含过多非生长活跃期,5°C在高纬又可能过于严格。
一个常被忽视的问题:这两个阈值是联动的还是独立的?假设一个像元的NDVI刚好在0.15–0.25之间摆动、温度在-2°C–3°C之间波动,那么NDVI > 0.2+温度 > 0°C的组合条件会让这个像元的有效数据量产生非线性坍缩。而这恰恰是温带春季过渡期的典型情况。
2.3 物候产品/算法动态确定:像素级别的精确,但引入新的不确定性
固定月份法和双阈值法本质上是在做"减法":从一个完整的12个月时间序列中切除若干不想要的片段。而物候算法则试图做"匹配":对每个像素、每年独立提取生长季的精确起止日期(SOS和EOS)。这是理论上最精确的方法,能逐像素捕捉空间异质性和年际变化。
目前在文献中出现的方法主要包括:
物候产品直接提取:Pang et al. (2025)在分析全球人工林对骤旱的响应时,采用MODIS MCD12Q2物候产品的Greenup和Dormancy日期,取多年平均值以减少年际波动。直接从权威数据产品提取,省时省力,但MCD12Q2在热带和干旱区的有效像元比例偏低。根据Yin et al. (2025)的评估,在全球1470个城市群中,MCD12Q2有效像元的比例仅在40–60%左右,热带城市的缺失尤其严重。Chen et al. (2025)使用TIMESAT软件拟合LAI时间序列提取SOS/EOS。TIMESAT需要预设上下包络线比例、适应强度等平滑参数,不同参数组合下SOS估计差异显著。
动态阈值法:Zhang et al. (2025)基于GLASS LAI,以振幅25%阈值定义SOS/EOS,同时将LAI > 2的像素视为全年生长季——这是对常绿植被的合理处理。Wu et al. (2022)对GIMMS NDVI3g采用三种算法(动态阈值NDVI_ratio=0.5、分段逻辑斯蒂、双逻辑斯蒂)取均值,通过多算法融合来降低单一算法偏差。
PELT变点检测法:Fang et al. (2023)在Scientific Data上发布的北半球光合物候数据集中,使用了一种相对新颖的方法:先用平滑样条过滤噪声和异常值(通过Savitzky-Golay滤波+Grubbs检验的20次迭代程序),再用PELT(Pruned Exact Linear Time)变点检测算法识别光合周期起止,最后以振幅10%、25%、50%定义不同严格程度的SOS/EOS。同时剔除峰值GPP < 25%最大峰值的周期。该数据集使用GOSIF-GPP而非传统植被指数,对常绿植被的光合物候识别优于NDVI/EVI法,与通量塔GPP的SOS/EOS相关性达0.78–0.80。Wang et al. (2024)则使用五参数Weibull函数拟合GPP年循环,并以GPP > 1 gC m⁻²持续360天以上为条件排除常绿站点。
曲率变化率法:Yin et al. (2025)指出,MCD12Q2和VNP22Q2两个主流物候产品在定义上存在本质差异:MCD12Q2使用EVI2振幅15%阈值,而VNP22Q2使用曲率变化率(CCR)的局部极值。这意味着同一像素的SOS在两组产品间可能相差2–4周。为减少误检测,Yin et al.在方法中限定了DOY搜索范围(北半球SOS: 30–150,EOS: 210–335;南半球反之),但MCD12Q2使用负DOY值表示前一年,需要额外转换。
物候算法面临的最大挑战并非算法本身,而是三个层面的误差传播:
第一,算法选择。不同物候提取算法之间的SOS差异可达2–4周,而这并非是某些算法比另一些"更正确":它们只是定义了不同的物候事件点(绿度起点 vs. 光合起点 vs. 冠层发育中点)。
第二,空间分辨率不匹配。MCD12Q2是500 m分辨率,但研究所用的气候数据和植被数据很可能在0.05°(~5.5 km)、0.1°或0.5°网格上。当你在粗网格上做空间平均时,一个格网内可能包含了物候期相差悬殊的不同像,平均后的结果可能不代表任何实际像元的情况。
第三,常绿植被的物候提取至今仍是难题。常绿类型的季节信号极弱,多数算法对其SOS/EOS的识别可靠性远低于落叶类型。Zhang et al. (2025)的LAI > 2方案和Wang et al. (2024)的全年GPP排除法都是在尝试处理这个问题。
2.4 生理阈值法:从"什么时候绿"到"什么时候在光合"
前面三类方法都在用植被指数(或其衍生的物候标签)来定义生长季,而生理阈值法放弃了这一路线,转而用纯环境变量定义"光合活跃期"。
Qi et al. (2024)的做法是:T > 15°C且VPD > 0.5 kPa且PPFD > 500 µmol m⁻² s⁻¹。这三个条件分别保证了温度适宜、蒸散发驱动力充足和光照充分,本质上定义的是一个"峰值生长季"(peak growing season),而非传统意义上的完整物候期。在峰值生长季,叶面积变化相对较小,GPP的异常更直接地归因于水分胁迫,而非物候驱动。
Fu et al. (2022)采用类似条件(T > 15°C、VPD > 0.5 kPa、SW > 250 W m⁻²),并通过"峰值3个月GPP均值"和"30天滑动平均距平法"两种替代定义做稳健性检验,结果一致。多重定义交叉验证是一种值得借鉴的做法。
Liu et al. (2025)在FLUXNET站点层面使用GPP百分位法:GPP > 年观测的30%分位数且TA > 0°C作为基本生长季,GPP > 70%分位数作为峰值生长季。30%的门槛比0°C+植被指数的双条件更为宽松(不预设固定的GPP绝对值),允许低生产力生态系统的数据进入分析,同时又排除了真正的休眠期。
生理阈值法的逻辑基础是:排除物候变化的干扰,只保留光合活性稳定的高峰期。这在分析水分胁迫对GPP的直接影响时非常合理——你不希望把"秋天自然落叶导致的GPP下降"和"干旱导致的GPP下降"混为一谈。
但它的核心局限同样明显:它定义的是峰值生长季而非完整生长季。春季萌发和秋季衰老这两个对干旱高度敏感的物候阶段会被排除。如果你的研究问题恰好涉及干旱对物候的直接影响(如干旱导致秋季叶片提前衰老),生理阈值法本质上预先排除了你试图观察的信号。
2.5 去趋势/距平法:绕过"筛"这个动作本身
最后一类方法在逻辑上最为激进,它干脆不筛。保留全年数据,用统计手段消除季节循环的影响,从而绕过生长季定义问题。
Vicente-Serrano et al. (2013)开创了这条路线:对每个像素计算12个月×24个时间尺度共288个SPEI-NDVI相关系数,取最大值作为该像素的干旱-植被响应强度。这种方法被誉为"不需要物候先验知识"的暴力统计,让数据自己说话,用最大相关系数所处的月份和尺度反推该像素的实际敏感时段。Zhang et al. (2017)、Li et al. (2024)沿用了此法。
Wang et al. (2025)则走双重去趋势路线:先去除长期线性趋势,再扣除多年月均值季节循环,以距平 < -0.5 SD定义干旱事件,同时用重心法(center of gravity)计算干旱事件的响应时间,而非简单的起止点差值,从事件整体角度更完整地刻画植被对干旱的响应过程。
去趋势/距平法的核心魅力在于避免了"在这一刀切"的主观判断,但它有一个隐蔽的前提假设:季节循环是稳定的、固定的,可以被干净地减除。这个假设在气候快速变化的背景下越来越不成立。春季提前、秋季推迟意味着季节循环本身在变形——你用历史平均来代表它,就会在变形的边缘产生残差。这些残差可能被误读为"干旱响应",实际只是物候漂移的统计伪影。
另一个问题是信噪比。低生产力区(如荒漠边缘带)的植被信号本身很弱,距平化后噪声/信号比进一步放大,极端事件的检出可靠性存疑。此外,距平法难以直接用于事件尺度的生态学分析。比如你想知道某次2012年北美大旱期间GPP损失了多少,距平法确实是会告诉你"那段时间距平是负的",但这无法等同于"完全由这次干旱事件造成的GPP损失"。
3 知识库文献统计:哪些方法最"流行"
从知识库收录的126篇生态遥感文献中的46篇明确定义了生长季筛选的研究来看,方法选择的分布并不均匀。
固定月份法(含纬度分区和VI掩膜) 以约15篇(33%)的论文数居首。对于大尺度分析,固定月份法在计算效率和可复现性方面无可匹敌。
仅温度/LAI/SIF单一阈值以约6篇(13%)紧随其后,VI+温度双条件法约5篇(11%)。这两类合计约24%,构成了第二大阵营。简单但比纯固定月份法多了一层生理约束。
物候产品/算法动态确定约8篇(17%)。研究者正在逐渐接纳逐像素动态判定的理念,但较高的技术门槛和数据质量不确定性限制了其普及速度。
去趋势/距平法约4篇(9%),生理阈值法约3篇(7%)。
另有约5篇(11%)没有明确说明生长季筛选方法。
从统计中还能提炼出几个有意义的信号:
第一,仅约41%的研究采用了逐像素动态判定(物候算法+生理阈值+去趋势法),其余要么使用固定规则,要么不做显式筛选。换言之,超过一半的研究在生长季定义上存在空间上的过度简化:它们在把温带落叶林、地中海灌丛和北方针叶林的物候期纳入同一个固定的月份框架,但很少在讨论部分充分承认这一假设的局限性。
第二,NDVI > 0.1是最常用的植被像素掩膜阈值(4篇),其次是NDVI > 0.2(3篇)。这两个临界值分别对应什么植被覆盖度、在不同生态系统中分别剔除多少数据,几乎没有论文做过细致的敏感性分析。
4 选择框架:基于研究问题的决策树
没有放之四海皆准的最优方法,但可以根据研究问题和数据条件做出有据可依的选择。以下尝试提供一个框架,供讨论和批判:
**如果你的分析是全球尺度、粗分辨率(≥0.5°),且目标是"年尺度上植被对气候的总体响应"**,那么纬度分区固定月份法(如O and Park 2024的三区方案)是一个可接受但必须完整报告的折中方案。务必在讨论中明确承认其对热带/亚热带的适用性有限。
如果你的分析需要精确的物候边界(如春季干旱对植被萌发时间的影响),则可以优先使用物候产品(MCD12Q2)或算法动态提取(TIMESAT/PELT/Weibull),并至少做一次算法间一致性检验。如果常绿植被占比大,考虑补充GPP或SIF作为判据,或参考Fang et al. (2023)的PELT+多振幅阈值框架。需要注意的是,不同物候产品的定义对齐(如MCD12Q2的15%振幅阈值 vs. VNP22Q2的曲率变化率)是一个隐蔽但影响巨大的系统偏差来源。
如果你的分析是站点级(如FLUXNET通量塔数据),且关心水分胁迫对光合的直接效应,生理阈值法(如Qi et al. 2024的T > 15°C+VPD > 0.5 kPa+PPFD > 500)或GPP百分位法(如Liu et al. 2025的30%/70%分位数)是逻辑一致的选择。建议对阈值本身做敏感性测试——15°C改为10°C、VPD阈值放宽至0.3 kPa,结果是否稳健?
如果你的分析重点是"气象-植被相关性",对事件时间定位精度要求不高,那么去趋势/距平法(如Vicente-Serrano 2013的最大相关系数法)提供了一种无需先验物候知识的替代方案。但需评估气候变化对季节循环形态的影响,并在讨论中考虑距平法对低信噪比像元的不确定性。
对所有研究而言,至少应当加一层NDVI > 0.1的掩膜来排除非植被信号,这是成本最低、收益最明确的操作。对于关心光合而非冠层绿度的研究,可以考虑用SIF阈值(如Li et al. 2023的SIF > 0.2 mW m⁻² sr⁻¹ nm⁻¹)替代或补充NDVI掩膜。
另一个常被忽视的维度是:你在用哪个指标定义生长季? NDVI反映的是冠层绿度(结构),SIF和GPP反映的是光合活性(功能)。这两种信号在大部分生态系统中高度相关,但在常绿森林和干旱胁迫期可能脱耦,因为植物可以在叶片尚绿时就已经关闭气孔、停止光合。因此,指标的选择应来自研究问题的推导,而非数据便利性。
最后,常绿植被需要单独处理。 Zhang et al. (2025)的LAI > 2(全年生长季)和Wang et al. (2024)的GPP全年排除法代表了两种截然不同的策略。前者将常绿植被纳入分析但假定了全年均匀生长,后者直接剔除以避免引入偏差。没有标准答案,但需要在论文中明确交代。
5 未尽的讨论
还有一些本文未能充分展开但值得在这个领域中持续关注的问题。
生长季定义的元问题:几乎所有方法都隐含了一个假设:生长季是一个可以被定义和识别的时间区间。但在气候变化背景下,物候边界本身在漂移。当你的研究对象(物候漂移)和你的筛选条件(固定月份)存在时空相关性时,筛选条件就不仅是预处理,而是分析框架的一部分,它可能同时排除并塑造你的结果。
多重证据链:单一方法永远有缺陷,但多个独立方法的交叉验证可以大幅提升结论的可靠性。例如:用固定月份法做基准分析,用物候产品做稳健性检验,在最关键的结论上要求两种方法方向一致。这一做法在Liu et al. (2025)的GPP > 30%+70%两层级方案和Fu et al. (2022)的多定义交叉验证中已有体现。
方法论透明性:无论选择哪种方法,在论文中完整报告就好:用了什么指标、什么阈值、逐像素还是固定规则、是否排除了常绿植被、阈值选择的依据是什么。Slette et al. (2019)曾专门发文指出"生态学家如何定义干旱以及我们为什么应该做得更好"是一个重要问题,类似的反省同样适用于生长季定义——一个方法的有效性,很大程度上取决于它是否可以被后来的研究者准确理解和复现。
论文信息
本文核心参考文献如下:
Deng, Y., et al. (2021). Responses of vegetation greenness and carbon cycle to extreme droughts in China. Agricultural and Forest Meteorology, 298–299, 108307.
Ding, Z., et al. (2020). Global analysis of time-lag and -accumulation effects of climate on vegetation growth. International Journal of Applied Earth Observation and Geoinformation, 92, 102179.
Fang, J., et al. (2023). Vegetation photosynthetic phenology dataset in northern terrestrial ecosystems. Scientific Data, 10, 300.
Fu, Z., et al. (2022). Atmospheric dryness reduces photosynthesis along a large range of soil water deficits. Nature Communications, 13, 989.
Ji, L., & Peters, A.J. (2003). Assessing vegetation response to drought in the northern Great Plains using vegetation and drought indices. Remote Sensing of Environment, 87, 85–98.
Konings, A.G., et al. (2017). Sensitivity of grassland productivity to aridity controlled by stomatal and xylem regulation. Nature Geoscience, 10, 284–288.
Li, W., et al. (2022). Widespread increasing vegetation sensitivity to soil moisture. Nature Communications, 13, 3959.
Li, W., et al. (2023). Widespread and complex drought effects on vegetation physiology inferred from space. Nature Communications, 14, 4640.
Li, D., et al. (2024). Declining coupling between vegetation and drought over the past three decades. Global Change Biology, 30, e17141.
Liang, S., et al. (2026). Vegetation in Central Asia is more sensitive to soil moisture stress. Agricultural and Forest Meteorology.
Liu, J., et al. (2025). When and where soil dryness matters to ecosystem photosynthesis. Nature Plants, 11, 1102–1112.
Lu, H., et al. (2026). Vegetation sensitivity shift from temperature to radiation dominance. Global Change Biology.
O, S., & Park, T. (2024). Global patterns of vegetation response to drought. Nature Communications.
Pang, J., et al. (2025). Flash droughts threaten global managed forests. Nature Communications.
Peng, S., et al. (2013). Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation. Nature, 501, 88–92.
Piao, S., et al. (2006). NDVI-based increase in growth of temperate grasslands and its responses to climate changes in China. Global Environmental Change, 16, 340–348.
Qi, G., et al. (2024). Soil moisture plays an increasingly important role in constraining vegetation productivity in China over the past two decades. Agricultural and Forest Meteorology, 356, 110193.
Vicente-Serrano, S.M., et al. (2013). Response of vegetation to drought time-scales across global land biomes. PNAS, 110, 52–57.
Wang, Y., et al. (2024). Increasing drought sensitivity of plant photosynthetic phenology and physiology. Global Change Biology.
Wang, Y., et al. (2025). Rising temperature increases the response time of LAI and GPP to meteorological drought in China. Atmospheric Research, 319, 107989.
Wang, X., et al. (2026). Climate warming reshapes vegetation responses in China from temperature to precipitation. GIScience & Remote Sensing.
Wen, Y., et al. (2019). NDVI indicated inter-seasonal non-uniform time-lag responses of terrestrial vegetation. International Journal of Applied Earth Observation and Geoinformation.
Wu, D., et al. (2015). Time-lag effects of global vegetation responses to climate change. Global Change Biology, 21, 3520–3531.
Wu, C., et al. (2022). Increased drought effects on the phenology of autumn leaf senescence. Nature Climate Change, 12, 943–949.
Xi, X., & Yuan, X. (2022). Significant water stress on gross primary productivity during flash droughts. Nature Communications.
Yin, P., et al. (2025). Comparative analysis of global urban land surface phenology between the MODIS and VIIRS products and extraction methods. Journal of Environmental Management, 375, 124326.
Zhang, L., et al. (2017). Response of vegetation to different time-scales drought across China. Remote Sensing, 9, 761.
Zhang, Y., et al. (2025). A pronounced decline in northern vegetation resistance to flash droughts. Nature Climate Change.
欢迎关注:疯狂学习GIS
本文参与 腾讯云自媒体同步曝光计划,分享自微信公众号。
原始发表:2026-06-01,如有侵权请联系 cloudcommunity@tencent.com 删除
评论
登录后参与评论
推荐阅读
目录
腾讯云开发者

Copyright © 2013 - 2026 Tencent Cloud. All Rights Reserved. 腾讯云 版权所有
深圳市腾讯计算机系统有限公司 ICP备案/许可证号:粤B2-20090059 ![]() 粤公网安备44030502008569号
粤公网安备44030502008569号
腾讯云计算(北京)有限责任公司 京ICP证150476号 | 京ICP备11018762号